As it is known, different cells have the similarity not only
in structure, but also in chemical composition as well, that indicates, in fact, that
their origin was subordinated to the common laws of the Evolution of Matter. The average
content of chemical elements in cells is the following (in percentage):
| oxygen | 65 - 75 |
| carbon | 15 - 18 |
| hydrogen | 8 - 10 |
| nitrogen | 1.5 - 3.0 |
| phosphorus | 0.2 - 1.0 |
| potassium | 0.15 - 0.4 |
| sulphur | 0.15 - 0.2 |
| chlorine | 0.05 - 0.1 |
|
|
| calcium | 0.04 - 2.0 |
| magnesium | 0.02 - 0.03 |
| sodium | 0.02 - 0.03 |
| ferrum | 0.01 - 0.015 |
| zinc | 0.0003 |
| cuprum | 0.0002 |
| iodine | 0.0001 |
| fluorine | 0.0001 |
|
From 104 elements of Mendeleev's periodical system more than
60 are found in cells. Atoms of oxygen, carbon, hydrogen and nitrogen fill in 98% of fnl.
cells of cellular subsystems. 1.9% are left to atoms of potassium, sulphur, phosphorus,
chlorine, magnesium, sodium, calcium and ferrum. Less than 0.1% of fnl. cells are occupied
by other substances (micro elements). Various combinations of the said elements give
several types of intracellular subsystemic formations, which every cell includes into
its fnl. cells as fng. units in the following proportions (in percentage):
| Inorganic |
| water | 70 - 80 |
inorganic
substances | 1.0 - 1.5 |
|
|
| Organic |
| proteins | 10 - 20 |
| fats | 1.0 - 5.0 |
| carbohydrates | 0.2 - 2.0 |
| nucleinous acids | 1.0 - 2.0 |
ATF and other low-
molecular
organic
substances | 0.1 - 0.5 |
|
All the above stated substances, being themselves very complex
in respect to the structure, are not piled up in a cell together in some chaotic disorder,
but as fng. units are filling in fnl. cells located in a strictly definite order and
destined for each of them in a uniform structure. While functioning they accomplish their
precisely defined micromotions inside a microvolume of a cell's space, regulated by
appropriate intracellular algorithms, at the same time there is an undoubted connection
of these motions in space with both the absolute and relative courses of time.
Each substance of a cell as a fng. unit carries out a strictly definite functional load
and has its own periods of functioning, regulated by appropriate algorithms. All their
various combinations constitute the unified, finely adjusted cellular mechanism.
Carbohydrates, fats and lipoids are attributed to the simplest
structural intracellular formations. Fnl. cells of their structures are being filled in
mainly by atoms of carbon, hydrogen and oxygen. The function of carbohydrates is
the most simple. Dissociating to CO2 and water, with emitting from each gram
4.2 large calories of energy, they supply with the essential mass of these fng. units
appropriate fnl. cells of the structure of cells.
The role of fatty compounds is more complicated. They add
to cells hydrophobias (waterproof) characteristics, and are heat-resistors. In the case of
necessity, they become, like carbohydrates, a source of accumulated energy, decomposing up
to CO2 and H2O. The dissociating of 1 gram gives 9.3 large calories.
Proteins are some more complex structural formations.
Besides carbon, hydrogen and oxygen in fnl. cells of their structures there are also
atoms of nitrogen, sulphur and other substances. Proteins are macromolecules combining
tens, hundreds of thousands of atoms. (So, if the molecular mass of benzol is equal 78,
then of protein of eggs is 36 000, of protein of muscles -
1 500 000, etc.)
The systemic organisation of proteins has its peculiarities.
Atoms entering into them fill in the fnl. cells destined for them not one by one, but by
the whole aminoacidic blocks, having a stable character of intrasystemic links. There are
altogether 20 of such fng. units - blocks. All of them have different systemic structures
and carry out different functions. Therefore the formation of proteins has a stage by
stage character.
At first aminoacids are being formed, which by means of peptidase
links are connected into proteinous chains with the giving off of water. Each proteinous
chain has on average of up to 200 - 300 aminoacidic blocks in different combinations. It
is enough to substitute in a chain one type of aminoacids for another one, as the entire
structure of a given protein, and with it its functional features as well are changing.
The structure of a proteinous chain of aminoacidic blocks has the form of a globule,
that adds to long chains of protein a compact appearance and mobility during spatial
displacements. In the packing of a polypeptidase chain there is nothing accidental or
chaotic, each protein has the definite, always constant character of packing. In other
words, the structure of every protein has a strictly definite spatial location of its
fnl. cells, which are being filled in by fng. units - aminoacidic blocks strictly
corresponding to them. At the same time each structure of protein, being a fng. unit in
a system of a higher order and occupying in it a fnl. cell corresponding to it, carries
out there its own function, characteristic only of it. As a rule, proteinous structures
are the most active reagents of chemical reactions, continually going inside cells, and
therefore their most important role is being catalysts of these reactions. Almost every
chemical reaction in cells is being catalysed by its own particular protein-ferment, the
catalytic activity of which is defined by a small part - its active centre (a combination
of aminoacidic radicals). The structure of a ferment's active centre and the structure
of a substratum precisely correspond to each other. They fit to each other as a key to
its lock. Because of the availability of a structural conformity between the active centre
of a ferment and substratum they can tightly approach each other, which actually provides
the possibility of a reaction between them.
To other important intracellular formations we should attribute
nucleinous acids: deoxyribonucleic - DNA and ribonucleic - RNA. Their main function
is to ensure the process of the synthesis of the cells' proteins. The length of a DNA's
molecule is a hundred and thousand times as big as the biggest proteinous molecule and can
reach several tens and hundreds of micrometers, while the length of the biggest proteinous
molecule does not exceed 0.1 mcm. The width of a DNA's double spiral is only 20
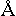 . The molecular
mass is tens and even hundreds of millions. Every DNA's chain is a polymer, monomers of
which are molecules of four types of nucleotides. In other words, DNA is a polynucleotide,
in the chain of which in a strictly definite order (and always constant for every DNA)
nucleotides are following, thus being fng. units in the structure of DNA's fnl. cells.
Therefore, if though in one of fnl. cells a different fng. unit - nucleotide is placed,
fnl. characteristics of the entire structure would change. In every DNA's chain (an
average molecular weight of 10 million) there are up to 30 thousand nucleotides (the
molecular weight of each being 345), owing to that the number of isomers (at 4 types
of nucleotides) is very great.
. The molecular
mass is tens and even hundreds of millions. Every DNA's chain is a polymer, monomers of
which are molecules of four types of nucleotides. In other words, DNA is a polynucleotide,
in the chain of which in a strictly definite order (and always constant for every DNA)
nucleotides are following, thus being fng. units in the structure of DNA's fnl. cells.
Therefore, if though in one of fnl. cells a different fng. unit - nucleotide is placed,
fnl. characteristics of the entire structure would change. In every DNA's chain (an
average molecular weight of 10 million) there are up to 30 thousand nucleotides (the
molecular weight of each being 345), owing to that the number of isomers (at 4 types
of nucleotides) is very great.
Because of the principle of complementarity as the basis of the
formation of a DNA's double spiral, a DNA's molecule is capable of redoubling. During this
process the two chains are separating, forming at the same time two double chains of fnl.
cells, only one row of which is filled in by fng. units, and the other one becomes free.
At the next stage dissociated nucleotides from the system's surroundings fill in free
fnl. cells which correspond to them in both spirals. As a result of the reduplication
in place of one molecule of DNA, two molecules originate of quite the same nucleotides'
composition, as the original one. One chain in each molecule of DNA originated anew is
left from the original molecule, the other one is being synthesised newly. In such a way,
together with the structure, the passing of fnl. characteristics of DNA from a motherly
cell to a daughter's one is occurring.
Graphically it looks like this:
The molecules of RNA are also polymers as are the DNA's,
but in contradistinction to them they have one spiral of fnl. cells and not two. RNA
carry out several functions in cells including:
1) the transport one (they are transporting
aminoacidic blocks to locations of the synthesis of proteins);
2) the informational one (they are transferring
the information about the structure of proteins);
3) the ribosomal one.
One more very important nucleotide in the structure of living
cells is adenosinthreephosphorous acid - ATPHA, the content of which in cells varies from
0.04 to 0.2 - 0.5%. Its peculiarity consists in the fact, that during a chipping off of
one molecule of phosphorous acid, ATPHA turns into ADP (adenosindiphosphorous acid) with
the emitting of 40 kilo joules of energy from 1 gr.-molecule.
All the above mentioned organic substances are complex in
their structure and in systemic organisation formations, but they in their also turn
enter as fng. units into fnl. subsystems of the cell's integrated system. To the
cell's basic subsystems the following ones are attributed:
The outward membrane of the cell. It is
regulating the entering of ions and molecules into the cell's structure and their leaving
it into the system's surroundings. Such an exchange of molecules and ions, that is of
different fng. units, between the cell's system and its surroundings is going continually.
One can distinguish the phagocyting, the taking up by the membrane of large particles of
a substance, and the pinocyting, the absorbing of water and water solutions. Through the
outward membrane the products of the cell's vital activity leave it, that is fng. units
having functioned in the cell's subsystems.
The cytoplasm. It is the internal semi-liquid
habitat of the cell, in the systemic volume of which the cell's internal structure is
expanded, that is its core, all organoids (or organelles), inclusions and vacuoles. The
cytoplasm consists of water with salts and various organic substances dissolved, among
which proteins predominate. The cytoplasm's structure consists of fng. units that are not
connected toughly but are moving freely along its entire volume. The fng. units filling
them in are transferred, when it is necessary, from them into the fnl. cells of organoids.
Therefore the cytoplasm's main functions are accumulative and transporting.
The endoplasmatic net. This is the cell's
organoid, constituting a complex system of canals and cavities, piercing the entire
cytoplasm of the cell. On membranes of the smooth endoplasmatic net the synthesis of
fats and carbohydrates takes place, which are being accumulated in accumulative fnl.
cells of its canals and cavities and then are being transported to different organoids
of the cell, where they occupy as fng. units appropriate fnl. cells of their structures.
On the membranes of canals and cavities there is also a great number of small rounded
bodies - ribosomes.
Each ribosome consists of two small particles, into the
composition of which proteins and RNA enter. Every cell has several thousand ribosomes
each. All proteins, entering into the composition of a given cell, are being synthesised
on ribosomes by means of the assembling of proteinous molecules from aminoacids, being
in the cytoplasm. The synthesis of proteins is a complex process of the filling in
with aminoacidic blocks of appropriate fnl. cells of their structures, which is being
accomplished simultaneously by a group of several tens of ribosomes, or by a polyribosome.
Synthesised proteins are being accumulated at first in the canals and cavities of the
granulated endoplasmatic net, and then are being transported towards those subsystems
of the cell, where fnl. cells destined for them are located. The endoplasmatic net
and polyribosomes constitute a single mechanism of biosynthesis, accumulation and
transportation of proteins.
The mitochondrias. This is an organoid,
the main function of which consists in the synthesis of ATPHA, representing a universal
source of energy, which is essential for the accomplishment of chemical processes
continually taking place inside the cell. The number of mitochondrias in the cell varies
from several to hundreds of thousands. Inside mitochondrias there are ribosomes and
nucleinous acids, and also a great quantity of various ferments. Synthesised ATPHA is
filling in transport fnl. cells of the cytoplasm and gets going towards the core and
organoids of the cell.
The plastids. They are organoids of
vegetable cells. They exist in several types. With the assistance of one of them,
chloroplasts, because of a pigment (chlorophyll) entering into their composition,
the cells of plants are capable of using the light energy of the Sun to synthesise
organic substances (carbohydrates) from inorganic ones. This process, as it is known,
has the name of photosynthesis.
The Golgy's complex. This is an organoid
of all vegetable and animal cells, in which the accumulation of proteins, fats and
carbohydrates takes place with their subsequent transportation to appropriate fnl.
cells both inside and outside the cell.
The lithesomes. This is an organoid, being
in all cells, that consists from a complex of ferments capable of breaking up proteins,
fats and carbohydrates. This is the main function of lithesomes. In every cell there are
tens of lithesomes, participating in the breaking up of already having functioned or
accumulative systemic formations as well as of those ones that get into the cell from
without by means of the phagocyting and pinocyting. As a result of breaking up fng. units
leave fnl. cells of being broken up structures, are being accumulated in fnl. cells of
accumulative systems of a given cell, and then are being transported to fnl. cells of its
new systemic formations. Having been broken up with the assistance of lithesomes, having
functioned the cell's structures are moved away out of its bounds. The formation of new
lithesomes takes place in the cell continually. The ferments, which are functioning in
lithesomes, as any other proteins are being synthesised on ribosomes of the cytoplasm.
Then these ferments get through the canals of the endoplasmatic net to a Golgy's complex,
in cavities and tubes of which fnl. cells of lithesomes' structures are being formed.
After being formed the lithesomes come off from tubes' ends and get into cytoplasm.
The cell's centre. This is an organoid,
which is located in one of parts of the concentrated cytoplasm. Two centrioles are
in it, which play an important role during the cell-fission.
The cell's structure has other organoids as well: flagellums,
cilias, etc., and also the cell's inclusions (carbohydrates, fats and proteins).
At the same time the cells, being themselves very complex
systemic formations, in their turn are fng. units, filling in fnl. cells of hypersystems
of the following levels of the organisation of Matter. Owing to this in the systemic
organisation of cells a mechanism is envisaged which allows within a relatively short
period of time the creation of systemic formations analogous to them. As a result the
cell's cycle includes two periods:
1) The cell-fission (a mitosis),
in the process of which two daughter cells are being created;
2) The period between two cell-fissions -
the interphase - the actual duration of a cell's functioning.
The cell's core plays an important role in the
cell-fission, being in every cell and constituting a complex fnl. subsystem. The core
has the core's membrane, through which proteins, carbohydrates, fats, nucleinous acids,
water and various ions get into and out of it. Having entered a core, they are filling
in fnl. cells of the core's juice as well as of nucleoluses and chromatin. In nucleoluses
the synthesis of RNA is taking place, but they themselves are being formed only in the
interphase. The chromatin constitutes a uniform substance, serving as an accumulative
subsystem, with the help of which the formation of chromosomes is being carried
out during the core-fission.
The chromosomes are the main mechanism of the cell, where so
named inherited information, which includes a chemical recording of the sequence of fnl.
cells in proteins' structures of a given cell, is being accumulated, kept and given
out. The above said information is being kept in DNA's molecules, which are situated
in chromosomes. Thus, DNA's molecules constitute a chemical recording of structures of
all the variety of proteins. On the lengthy thread of a DNA's molecule a recording of
information about the sequence of fnl. cells of various proteins' structures is following
one after another. A part of DNA, having the information about the structure of a protein,
it is usual to name a gene. A DNA's molecule constitutes a collection of several
hundreds or thousands of genes. The diameter of chromosomes is not big and amounts on
average to 140 , their
length, repeating the length of DNA's molecules, can be more than 1 mm. In the middle of
the interphase period the synthesis of DNA occurs, as a result of which a chromosome is
doubling.
The most important function of chromosomes is to be a repository
of the recordings of structures and accordingly of algorithmic abilities of the cell's fnl.
subsystems with the assistance of the mechanism of formation of proteinous fng. units.
In the course of time as functions of this or that type of organic systems are
increasing, the recording in chromosomes is changing and perfecting itself, meeting
the requirements of laws of the fnl. development of Matter. In a direct dependence
on a molecular recording of chromosomes' DNA through the mechanism of synthesising of
proteinous molecules, all the processes of vital activity of cells are occurring. The
number of chromosomes is constant for each species of animals and plants, that is each
cell of any organism which belongs to the same species contains an absolutely definite
number of chromosomes (rye - 14, man - 46, hen - 78, etc.). The chromosomes' composition,
which the core of a cell contains, always has twin chromosomes. So 46 chromosomes of
a man form 23 pairs, in each of them two identical chromosomes are united. Chromosomes
of different pairs differ from each other in form and place of location. As a result of
mitosis two daughter cells are being created, which by structure are fully similar to a
mother one. Each of them has exactly the same chromosomes and the same number of them as
the mother cell. In this way a complete communication of all the inherited information
to each of the daughter cores is provided. The core and all the organoids of a cell's
cytoplasm are interacting as a single system.
All cells have a similar type of the structure: the core,
mitochondrias, the Golgy's complex, the endoplasmatic net, ribosomes and other organoids.
However, before becoming such a perfect system, which it is nowadays, the cell has passed
a long way through the evolution, marked by appropriate spaces on ordinates of t and
ft of the tensor of the Evolution of Matter. In the beginning it was a part
of non-cellular organisms unknown to us, then of imperfect unicellular and multi-cellular
organisms, including bacteria and blue-green algae, and finally it reached the perfection
of a complex cellular mechanism, characteristic of the representatives of the vegetable
and animal world contemporary with us. Because of the motion of Matter along the ordinate
of quality during the process of the evolution of the cell a great variety of its
types was originated, each of them was provided with strictly definite fnl. features and
correspond to the definite point on this ordinate.
At the same time from a certain moment this process started
going simultaneously with the beginning of the development of fnl. systems of a higher
organisational level, fnl. cells of which the cells began to fill in as fng. units. As
a result the cell turned into a complex systemic formation, to keep up fnl. features of
which complex chemical processes are taking place continually inside and outside it. The
permanency of processes is connected with the fact that the time of the functioning of
fng. units with the growth of their molecular weight does not coincide more and more with
the time of the existence of fnl. cells of structures, that they fill in, as in a limited
space of displacement of fng. units the time of their existence is in direct dependence
on their fnl. mass. Besides, the permanency of processes is caused by the fact that most
chemical reactions taking place in a cell have an irreversible character. For all these
reactions the greatest organisation and order are characteristic: each reaction is going
at a strictly definite place at a strictly definite time in a strictly definite sequence.
Molecules of ferments are located on membranes of mitochondrias and of the endoplasmatic
net in the order in which reactions are going.
In a cell there are about one thousand ferments, with the
assistance of which two types of reactions are going: of synthesis and of
desintegration. As the main (creating) type of reactions should be considered
reactions of synthesis, in the process of which complex molecular compounds are being
formed, as fng. units filling in fnl. cells of the cell's subsystemic structures. So,
for replacement of each functioned out molecule of protein, that has left this or that
fnl. cell, a new molecule of protein fills the vacated place, by structure and chemical
composition and accordingly by its fnl. features fully identical to the previous fng.
unit. It means, that a newly synthesised fng. unit is able (or should be able) to take
an identical part in any algorithms, characteristic for a given fnl. cell.
The synthesis of fng. units is carried out with the assistance
of the functioning of the cell's special subsystems on the basis of the coded gene
recording of DNA. Fluctuatal deviations, which happen during this, in case of their
positive effect are being recorded by the reverse connection in a gene recording and
serve to the purposes of a further perfection of a given systemic structure. In the
event of a negative effect from a newly synthesised fng. unit the implementation of
a part of fnl. algorithms is being violated and in case the system is not able to
eliminate that, the unproper functioning of an appropriate subsystem can result in the
end in the destruction of the structure of a given cell as a whole. In this way the
cell's organisational system permits it to keep up a permanent presence of appropriate
fng. units in fnl. cells of their subsystems, that keeps its structure and by what the
cell's ability to implement algorithms of fnl. cells of systems of a higher order is
provided, where it enters as a fng. macro unit. All reactions of biosynthesis (reactions
of assimilation) take place according to the general theory of systems by absorbing
energy of motion in space, which as if getting stuck in the structure of the cell's
system is being transformed into energy of connection between its fng. units.
The other type of reactions - reactions of desintegration -
takes place with a simultaneous decrease in the energy of connection, being transformed
into energy of motion in space. During reactions of dissimilation, fng. units of the
cell's subsystems, being systemic formations of a lower order, having functioned out,
decompose to fng. units of their sublevel, ready if necessary to enter into new
synthesising reactions in order to form new structures - fng. units of a higher
organisational level. Both types of reactions are closely interconnected and constitute
a single process, directed to filling in fnl. cells of the cell's structure with active
appropriate fng. units, which finally provides the maintenance at a proper level of fnl.
features of the cell as a whole.
One of the main and the most complex types of synthesising
reactions is biosynthesis of proteins, taking place in the cell continually
during the entire duration of its existence. During the process of functioning of the
cell a part of its proteins, having participated in catalytic reactions, are being
denatured gradually, their structure and consequently their functions are being violated
and they are being moved away from their fnl. cells and then from the cell itself. Their
places in fnl. cells are being occupied by newly synthesised proteinous molecules
completely identical by its fnl. features to fng. units having emptied places for them.
Taking into consideration that there are a great number of types of proteinous molecules,
the mechanism of their synthesising, being perfected during a long period of time, in the
end turned into a specialised subsystem of the cell with the precise list of algorithms
of functioning.
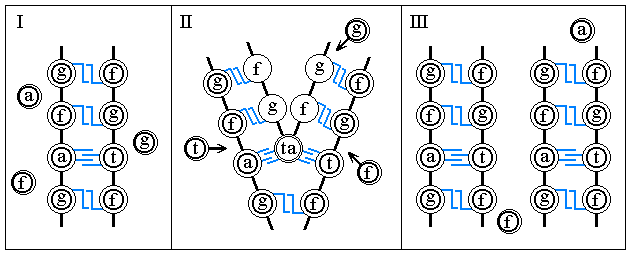
The program of synthesis of proteins, that is the information
about their structure, recorded and kept in DNA, is sent to ribosomes with the help of
informational RNA (i-RNA), being synthesised on DNA and precisely copying its structure.
To each aminoacid a section of a DNA's chain corresponds from three nucleotides being
situated alongside: A-C-A (cysteine), T-T-T (lysine), A-A-C (leucine), etc. The number
of possible combinations from 4 nucleotides by 3 equals 64, though in all 20 aminoacids
are used. The sequence of nucleotides of an i-RNA repeats precisely the sequence of
nucleotides of one of chains of gene recording, while from each gene it is possible
to make any number of copies of RNA.
 The recording of information on an RNA, that is the process of 'transcription', takes
place during the simultaneous synthesising of an i-RNA, which is being carried out with
the help of the principle of complementation. As a result, the chain of an i-RNA being
formed by content and sequence of its nucleotides constitutes a precise copy of the
content and sequence of nucleotides of one of the chains of DNA. The molecules of an
i-RNA are directed then to ribosomes, where aminoacids also come, being delivered from
without of the cell in already ready form. Aminoacids get to a ribosome accompanied by
transport RNAs (t-RNA), consisting on average of 70 - 80 nucletidic links, in 4 - 7
places complemented to each other. To one of a t-RNA's ends an aminoacid is being
connected and in the upper part of the bend a triplet of nucleotides is fixed, which
by code is corresponding to a given aminoacid. For every aminoacid there is its own
t-RNA, that is there are also 20 varieties of them.
The recording of information on an RNA, that is the process of 'transcription', takes
place during the simultaneous synthesising of an i-RNA, which is being carried out with
the help of the principle of complementation. As a result, the chain of an i-RNA being
formed by content and sequence of its nucleotides constitutes a precise copy of the
content and sequence of nucleotides of one of the chains of DNA. The molecules of an
i-RNA are directed then to ribosomes, where aminoacids also come, being delivered from
without of the cell in already ready form. Aminoacids get to a ribosome accompanied by
transport RNAs (t-RNA), consisting on average of 70 - 80 nucletidic links, in 4 - 7
places complemented to each other. To one of a t-RNA's ends an aminoacid is being
connected and in the upper part of the bend a triplet of nucleotides is fixed, which
by code is corresponding to a given aminoacid. For every aminoacid there is its own
t-RNA, that is there are also 20 varieties of them.
The synthesis of proteins and of nucleinous acids takes place
on the basis of reactions of matrix synthesis. By that the giving of fnl. features of
fng. units being replaced by newly formed compounds is provided. New molecules are being
synthesised in precise correspondence with the plan, which is kept put in the structure
of already existing molecules. Therefore in these reactions a precise, strictly specific
sequence of monomeric links in polymers that are being synthesised is provided. What is
taking place here is a directed pulling together of monomers to a certain place of the
cell - into fnl. cells of a being newly formed polymer, while the location of fnl. cells
themselves is being pre-determined by the structural organisation of a matrix being copied.
Macromolecules of nucleinous acids of DNA and RNA are playing the role of a matrix in
matrix reactions. Monomeric molecules (nucleotides or aminoacids) in accordance with the
principle of complementation are being located and fixed on the matrix in a strictly
definite, given order. Then a 'sewing together' of monomeric links into a polymeric chain
takes place, and a ready polymer is released by the matrix. After that the matrix is ready
for the assembling of a new polymeric molecule. With the help of a matrix type of reactions
the reproduction of the same type compounds - fng. units of a given system - is being
carried out. The necessity of the reproduction of the same type of fng. units is traced
through all levels of the organisation of Matter and is one of the main regularities of
the general theory of systems.
The information about the structure of a protein, recorded
on an i-RNA as a sequence of nucleotides, is being transferred further as a sequence
of aminoacids into a polypeptidase chain being synthesised, that is the process of
'translation' is taking place. During the assembling of a proteinous molecule, a
ribosome creeps along an i-RNA, after it the second one, then the third, etc. Each
of them synthesises quite the same protein, programmed on a given i-RNA. When the
ribosome passes along an i-RNA from one end to the other - the synthesis of a protein
is over. After that the ribosome goes on to another i-RNA and the protein is directed
through the endoplasmatic net into a free fnl. cell with features that correspond to
it, which it fills in as a fng. unit.
The synthesis of proteins in a cell takes place continuously.
All the ribosomes located simultaneously on one i-RNA are united into a polyribosome.
The ribosome works along an i-RNA taking 'short steps': triplet after triplet the RNA
is in contact with it. For the sewing of a polypeptidase chain in the ribosome there is
the protein-synthethasa. Molecules of a t-RNA, passing through a ribosome, touch by its
codic end the place of contact of the ribosome with an i-RNA. If a codic triplet of the
t-RNA turns out to be complementary to a triplet of the i-RNA, an aminoacid delivered
by the t-RNA moves over from its fnl. cell into a fnl. cell of a molecule of a protein
that is being synthesised, thus becoming a fng. unit of its structure. By this the most
important rule of the construction of fnl. systems is provided - the placing of a given
fng. unit into a fnl. cell strictly corresponding to it or, on the contrary, the filling
in of a fnl. cell with a fng. unit strictly corresponding to it. Therefore, the mechanism
of the synthesis of proteins, being available in any cell, provides a full guarantee
that a given aminoacid, being transported by a t-RNA, will get only into a fnl. cell
corresponding to it of a protein's structure or, on the contrary, that into a coming
up on the ribosome next in turn empty fnl. cell of a protein being synthesised only
a fng. unit - a required aminoacid corresponding to it by its fnl. features - will get.
After the filling in of a fnl. cell next in turn of a
synthesised protein, the ribosome is making one more step along the i-RNA, getting this
way the information about fnl. features of a fnl. cell which is next in turn in a being
filled structure. The t-RNA with the vacated working t-fnl. cell leaves into the
intracellular space, where it takes a new molecule of aminoacid corresponding to it in
order to carry it again to any of the fng. ribosomes. The molecules of proteins are
synthesised on average in about 1 - 2 minutes. This process takes place during the whole
period of a cell's existence. All the reactions of the synthesis of proteins are being
catalysed by special ferments, up to reactions of seizure by t-RNAs. All the reactions
of synthesis are endothermic and therefore each phase of the biosynthesis is always
linked with consumption of ATPHA.
Any cell keeps its composition and all its fnl. features at a
relatively constant level. So the content of ATPHA in cells is 0.04% and this magnitude
is kept stable. The starting and ending of processes, providing the keeping up of fnl.
features of a cell, happen in it automatically. The basis of auto regulation of these
processes is a special signal subsystem of cells, which uses for these purposes the fnl.
features of fng. units of previous sublevels, that is electromagnetic characteristics
of electrons, atoms, etc. Therefore in any cell there is always a certain quantity of
various ions and other charged particles, which as a whole creates bioelectrical chains,
microfields, etc. An alteration of the bioelectrical potential though in one of links of
any subsystem of a cell serves as the signal for the beginning or ending of this or that
biochemical reaction, of this or that transference of fng. units along fnl. cells of
various subsystems of the cell. The availability of the subsystem of signal bioelectrical
connection in the structure of cells as well as using for these purpos